Миграции ариев от 6000 до 3000 лет назад (часть 4)
В предыдущих трех частях этой серии статей (часть 1, часть 2, часть 3) рассказывалось про продолжительную миграцию предков ариев, носителей гаплогруппы R1a, из Южной Сибири по «южной дуге» через Анатолию (Малую Азию) с приходом на Балканы примерно 10-9 тысяч лет назад, предположительное создание ими культуры Лепенский Вир (9400-8200 лет назад), на что указывает положение костяков в этой культуре, характерное для R1a, начало отсчета периода существования исторических ариев от гаплогруппы-субклада R1a-Z645 примерно 5500 лет назад, которое также маркирует расхождение праиндоевропейских языков на ветви (субклады ариев Z93, Z283-Z282-Z280, M458, Z284, которые образовались, по данным геномного анализа и ДНК-генеалогии, 5000±500, 5000±500, 4700±500 и 4500±700 лет назад, соответственно), последующий переход ариев (носителей гаплогруппы R1a) по Русской равнине на восток, с образованием цепи их археологических культур – трипольской, шнуровой керамики, фатьяновской, катакомбной, бабинской, потаповской, срубной, петровской, андроновской, синташтинской, карасукской, тагарской, бактрийско-маргианского археологического комплекса (БМАК) и других культур, маркирующих миграционные пути исторических ариев.
http://pereformat.ru/wp-content/uploads/2016/08/r1a-migration28.jpg
Было показано, кто являются современными потомками этих культур, в частности, современные этнические русские, украинцы, белорусы, поляки, таджики, узбеки, туркмены, пуштуны и другие народы (в их части, имеющей гаплогруппу R1a), и показаны ДНК-генеалогические свидетельства перехода ариев в Индию, в Митанни, обсуждались хетты как наиболее вероятные потомки ариев. В этой, четвертой части, рассказывается о переходе ариев на Иранское плато, и продолжении более северных арийских миграций в Зауралье, на Алтай, в Китай, и о сопряженных с этим загадках про афанасьевскую культуру, Таримский бассейн, носителей тохарских языков. На этом рассмотрение материала заканчивает с восточными миграциями ариев, и в последующих частях переходит к миграциям ариев гаплогруппы R1a в связке фатьяновской культуры с западными миграциями с формированием балтийских славян, начиная с лужицкой культуры в конце II тыс. до н.э., карпатских ветвей гаплогруппы R1a и распространением их на Балканы, в Малую Азию, с образованием венетов-венедов и последующим распространением потомков ариев по Европе.
Переход ариев в Иран и «арии Авесты».
Первые три (не обязательно в хронологическом порядке) ветви арийских миграций были описаны в предыдущей, третьей части этой серии статей. Переходим к четвертой ветви. Из текстов «Авесты» следует, что арии перешли на Иранское плато из Средней Азии, возможно, с гор Таджикистана, и по оценкам переход состоялся примерно 3500 лет назад, после жизни в горах Средней Азии в течение примерно 500 лет. Хронологическая картина складывается в следующем виде – примерно 4500 лет назад арии начали расходиться с Русской равнины, примерно 4000 лет назад они достигли своих основных «перевалочных пунктов» – гор Кавказа на пути в Малую Азию и на Ближний Восток (чтобы стать хеттами и митаннийскими ариями), гор Средней Азии для последующего пути на Иранское плато (чтобы стать «авестийскими ариями» и далее персами и иранцами), и Южного Урала (и БМАК) для последующего перехода в Индию. Все эти дальнейшие переходы с «перевалочных пунктов» проходили с исторической точки зрения довольно синхронно, около 3600-3500 лет назад. Из этой хронологической картины становится понятно, почему в Авесте не отражены территории исхода ариев с Русской равнины, кроме зыбких воспоминаний об «арийском просторе», airyanəm vaējah (на авестийском языке). Ясно, что 500 лет пребывания в отрыве от предыдущих мест обитания практически стерли историческую память ариев.
Теперь, когда «арийские» вехи расставлены, перейдем к иранским гаплотипам. Поскольку достаточно общим положением является то, что арийские миграции в Индостан и на Иранское плато произошли примерно в одни и те же времена, мы можем ожидать, что и ДНК-генеалогия покажет те же самые датировки общих предков гаплогруппы R1a в Миттанни (и последующие гаплотипы гаплогруппы R1a у арабов), Индии и Иране. Как было показано в наших ранних исследованиях, арабские гаплотипы гаплогруппы R1a происходят от общего предка (или общих предков), которые жили примерно 4050±400 лет назад, индийские гаплотипы – от общих предков (в основном субкладов L657 и Z2123), которые жили 4600±485 лет назад.
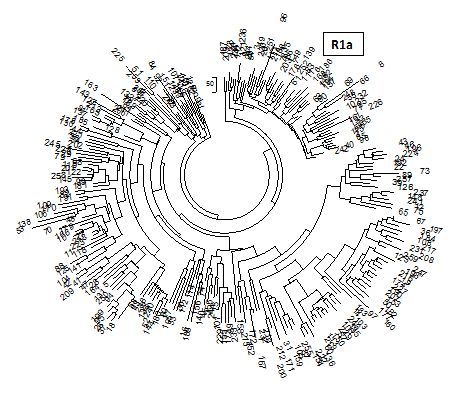
Посмотрим на иранские гаплотипы. Протяженных иранских гаплотипов в коммерческих базах данных почти нет (в базе данных IRAKAZ есть два 67-маркерных гаплотипа), поэтому обратимся к более коротким гаплотипам из полевых выборок. Не так давно опубликована статья иранских популяционных генетиков, в которой приведены 259 восточно-иранских гаплотипов в 17-маркерном формате (Hedjazi et al., 2014). Поскольку это статья популяционных генетиков, то в статье, конечно, нет никакой ДНК-генеалогии, но один плюс перевешивает – там приведены гаплотипы. В остальном – гаплогруппы не определяли, все гаплотипы свалены в одну кучу, авторы сообщили, что из 259 гаплотипов 256 «уникальны», то есть встречаются только по одному разу, а три гаплотипа встречаются дважды, потому индекс «разнообразия гаплотипов» равен 0.9999. Как мы не раз отмечали, это совершенно бессмысленные показатели, и никому эти «разнообразия» никогда не были нужны. Они приводятся в статье по трем регионам восточного Ирана, опять же без разделения на гаплогруппы, опять на уровне «разнообразия» и без какого-либо обсуждения. Вот, собственно, и всё. Это – показательная популяционная генетика. Такая у них наука. Ладно, спасибо за гаплотипы, хотя разделение их на гаплогруппы пришлось делать самим. Дерево гаплотипов приведено на следующем рисунке (естественно, в цитируемой статье его не было).
Дерево 259 иранских гаплотипов Y-хромосомы в 17-маркерном формате. Справа вверху – ветвь из 58 гаплотипов гаплогруппы R1a. Построено по гаплотипам, представленным в приложении к статье (Hedjazi et al, 2014). Общий предок ветви R1a жил 4900±600 лет назад; остальные ветви – серии древних гаплотипов гаплогрупп (по часовой стрелке) J2, L1a, L1c, G2a, R1b, Q, J1, J2, R2a, E1b (повторы гаплогрупп – это отдельные ветви этих гаплогрупп). Общие предки древних ветвей жили 4-8 тысяч лет назад. Это, видимо, древние доарийские племена лулубеев, касситов, гутиев, хурритов, эламитов, а также потомки относительно недавних визитеров в Иран.
Базовый гаплотип ветви восточно-иранских R1a на дереве выше:
13 25 16 11 11 14 10 13 11 31 – 16 14 20 12 15 11 23
Он отличается от базового индийского гаплотипа R1a на две мутации (отмечено)
13 25 16 10 11 14 10 13 11 30 – 16 14 20 12 15 11 23
и от базового гаплотипа этнических русских на три мутации
13 25 16 11 11 14 10 13 11 30 – 15 14 20 12 16 11 23
На самом деле, поскольку усредненные мутации дробные, различия составляют всего 1.26 и 1.62 мутаций, соответственно, то есть разница между общими предками восточно-иранских и индийских гаплотипов составляет 900 лет, а восточно-иранских и праславянских (R1a) гаплотипов – 1150 лет, и их общие предки жили, соответственно, примерно 5200 и 5500 лет назад, что практически совпадает в пределах погрешности расчетов, и согласуется с датировками субкладов, приведенными выше. Это всё арийские общие предки субклада R1a-Z645. Еще один вывод, который следует из приведенных расчетов, что иранские и индийские арийские носители гаплогруппы R1a разошлись c разницей примерно на 450 лет, что не противоречит нашим представлениям о разнице в их языке, пантеоне богов, и в целом о динамике миграций ариев раздельно в Индию и Иран.
Эти миграции были частично рассмотрены в предыдущих частях данной серии статей, в разделе о синташтинской и андроновской культурах, которые затем переходят в карасукскую (3700-2900 лет назад) культуру Южной Сибири и Казахстана, которая затем сменяется тагарской (2800-2300 лет назад) и таштыкской (2200-1500 лет назад) культурами. Ископаемые ДНК карасукской культуры показали (Allentoft et al., 2015) гаплогруппу R1a-Z2124 (захоронение Арбан-1 в Минусинской котловине, без датировки, нижестоящие Z2122 и Z2123 отрицательные) и R1a-Z645 (3340±70 лет назад, захоронение Сабинка 2). В тагарской культуре все ископаемые ДНК показали (Keyser et al., 2009) гаплогруппу R1a, с археологической датировкой 2800-1900 лет назад. В межовской культуре (Капова пещера, северная часть Челябинской области) найдена ископаемая гаплогруппа R1a-M417, там типирование скорее всего поверхностное, неглубокое, субклад М417 образовался 8500±1300 лет назад, по данным коллектива YFull на основании геномных данных. Сама межовская культура датируется 3300-2900 лет назад.
Уйдя за Южный Урал, оставив после себя часть андроновской археологической культуры северного Казахстана, срубную, потаповскую, синташтинскую культуру и им родственные, арии и здесь разошлись на миграционные потоки. Часть их направились на юг, в итоге прибыв в Индию, другие стали продвигаться дальше на восток, причем скорость их продвижения была поразительной – за несколько столетий они продвинулись на несколько тысяч километров, дойдя до Хакасско-Минусинской котловины. Именно там были найдены ископаемые R1a с датировкой 3800-3400 лет назад (рис. ниже). Это – немногим более поздние, чем в синташтинской культуре R1a, как и должно быть (те были Z645 c датировкой 4030±115 лет назад, и М420 c датировкой 4190±130 лет назад). Это всего лишь тысячу лет спустя от находки ископаемой гаплогруппы R1a в Германии (4600 лет назад). Правда, находка в Германии наиболее вероятно относится к субкладу Z645-Z280, а в Абакане-Минусинске к Z645-Z93, в таком случае они происходят не один из другого, а от общих предков, субклад которых был Z645, но все равно скорость миграции носителей R1a на восток впечатляет.
http://pereformat.ru/wp-content/uploads/2016/08/r1a-migration30.jpg
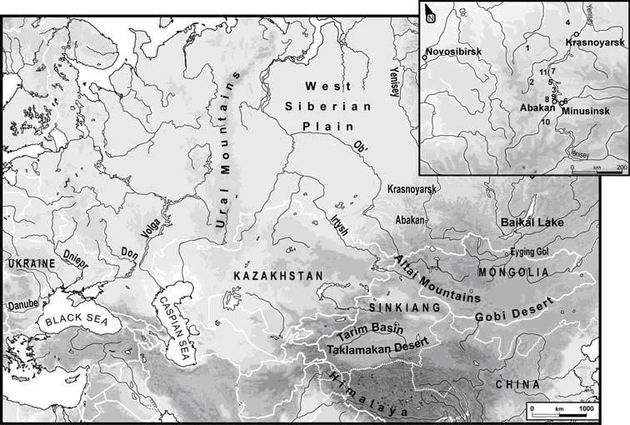
Ископаемые R1a с археологической датировкой 3800-3400 лет назад были найдены в районе Абакана-Минусинска, за несколько тысяч километров на восток от Урала, к северу от монгольской границы, в приалтайском регионе. На врезке показаны (нумерация) расположения археологических участков, из которых брали костные материалы для определения ДНК (Keyser et al., 2009).
Получается, что примерно во времена захоронений ариев в далеком Зауралье, на Алтае, их другая группа уже прибывала в Индию, а также на Иранское плато.
Ископаемые гаплотипы гаплогруппы R1a (3800-3400 лет назад на Алтае):
13 25 16 11 11 14 10 14 11 32 – 15 14 20 12 16 11 23 (древние R1a, андроновцы)
На той же территории находятся захоронения тагарской археологической культуры, и в них тоже найдены ископаемые R1a, c датировками 2800-1900 лет назад. Хотя прошли одна-две тысячи лет, гаплотипы отстались почти такими же:
13 24/25 16 11 11 14 10 13/14 11 31 – 15 14 20 12/13 16 11 23 (древние R1a, тагарцы)
В этом и состоит надежность гаплотипов – они меняются незначительно с течением времени. Если они изменились по нескольким маркерам – следовательно, прошли тысячелетия. В последующей за тагарской, таштыкской культуре (2200-1500 лет назад) тоже найдены ископаемые ДНК гаплогруппы R1a, гаплотипы которых почти идентичны тагарским, с отличием всего на одну мутацию.
После рассмотрения этого материала, и перед тем, как продолжить, вернемся к определениям. Как было указано в первой части настоящей серии, ариев можно определить от начала образования гаплогруппы R1a-Z645, примерно 5500 лет назад, и далее по совокупности нижестоящих субкладов, вплоть до середины I тыс. до н.э., поскольку термин «арии» тогда еще встречался помимо Индии, а именно на территории, которая впоследствии будет названа Персией, а затем Ираном. Тогда это определение согласуется с временем начала расхождения арийских (индоевропейских) языков на ветви, с миграциями ариев на юг (хетты и митаннийские арии), юго-восток (авестийские арии), на восток (индоарии, китайские арии и «скифский круг» археологических культур на Алтае). Это определение вполне конкретно, и практически ничего не нарушает в парадигмах историков и лингвистов. Тогда этнические русские, украинцы, белорусы, поляки, сербы, хорваты, словенцы, чехи, словаки, греки и другие европейцы гаплогруппы R1a (кроме разве что северо-западных европейцев гаплогруппы R1a-L664), а также носители гаплогруппы R1a среди башкир, татар, карачаево-балкарцев, таджиков, узбеков, пуштунов, уйгуров и многих других народов Европы и Азии являются потомками ариев. Ничего плохого или предосудительного в таком определении нет, народы имеют право знать свою историю. Языки у них индоевропейские, тюркские, финно-угорские, и опять же ничего в этом неправильного нет. Языки меняются, гаплогруппы остаются, субклады со временем ветвятся. Нормальный путь эволюции человека. Вот так определение ископаемых гаплотипов продвигает наши знания, конечно же в совокупности с обширными данными ДНК-генеалогии и других дисциплин.
Дальнейшее продвижение носителей гаплогруппы R1a от Урала на юг и восток примерно 4000 лет назад привело к очередным загадкам современной науки. Ранее было рассказано, что гаплогруппа R1a образовалась примерно 20 тысяч лет назад, видимо, в Южной Сибири, хотя прямых доказательств, в виде ископаемых гаплотипов, этому пока нет. Но если это так, то по мере продвижения туда через многие тысячелетия носителей R1a из Европы, в основном, как показывают исследования ископаемых гаплотипов, субкладов Z645-Z93 (и нижестоящих, как Z2123), они могли перемешиваться с автохтонными (неевропейскими по происхождению) гаплогруппами R1a, субклады которых нам могут быть пока неизвестны. Если субклады окажутся Z645-Z280, то это европейские по происхождению субклады, они сейчас доминируют у славян гаплогруппы R1a. Вполне возможно, что они тоже в древности прошли в Зауралье, на Алтай и в Иран-Индию-Месопотамию, просто их там пока не нашли при археологических раскопках. Но если на Алтае, в Монголии и в Китае окажутся субклады R1a (xZ645) [то есть R1a, кроме Z645], то есть в упрощенном виде R1a (x Z93, Z280), то это прямой сигнал к тому, что мы нашли автохтонные R1a. Тем более если они окажутся древнее 4000 лет назад. Поэтому, изучая гаплогруппу R1a в тех краях, нужно постоянно иметь это в виду, а именно, что R1a могут иметь там совершенно разную историю – автохтонные и пришлые.
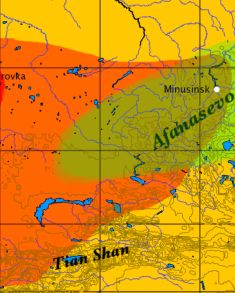
Расположение афанасьевской культуры на карте. Слева – расположение восточной части андроновской культуры. Афанасьевскую культуру часто связывают с Алтаем и Хакассией, но в нее включают также Восточный Казахстан, Западную Монголию, и Синцзянь на северо-западе Китая.
На Алтае есть загадочная афанасьевская культура. Загадочная она по разным причинам. Несмотря на то, что первый ее могильник был исследован еще в 1920 году, до недавнего времени справочники и учебники не указывали возраст этой культуры. Сейчас она обычно датируется III-II тыс. до н.э., другие источники дают 5200-4500 лет назад. Далее, современные источники совершенно запутаны в отношении происхождения этой культуры. Одни историки пишут, что она была основана «мигрантами из Восточной Европы», а именно «носителями древнеямной культуры». На самом деле носители древнеямной культуры прибыли в Европейскую часть современной России как раз из Южной Сибири, и «мигрантами из Восточной Европы» их в этом контексте назвать никак нельзя. Другие историки пишут, что «наследниками афанасьевцев были племена тагарской культуры», но эти никак не совместимо с ямниками как создателями афанасьевской культуры, поскольку ямники – это гаплогруппа R1b, а тагарцы – R1a. Пишут, что афанасьевская культура сменилась карасукской, но это тоже несовместимо с ямной культурой как относящейся к гаплогруппе R1b, и карасукской гаплогруппы R1a. Пишут, что «афанасьевская культура ассоциируется с прото-тохарами, потомков которых, тохаров, скифы вытеснили в китайский Синцзянь». Это вообще ни на чем не основано, так, «по понятиям». Собственно, глухое слово «ассоциируется» на это и указывает.
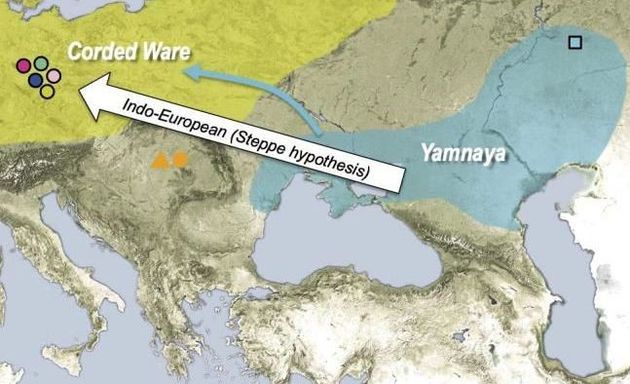
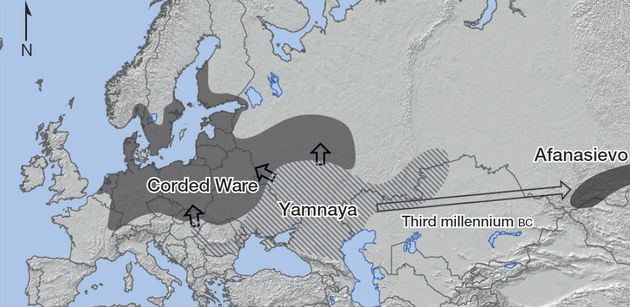
Оставим в стороне рассмотрение многоходовых фантазий, но чуть позже покажем, что в Синцьзяне нашли только ископаемые R1a, то есть опять ямники здесь не причем. Но ямники (R1b) упорно держатся усилиями теперь генетиков как предки R1a, что, конечно, абсурд. Совсем недавняя работа по изучению генома ископаемых ДНК (Allentoft et al., 2015) мало того, что объявила, что культура шнуровой керамики (R1a) произошла от ямной (R1b), но и то, что афанасьевская культура также произошла от ямной (см. их представления на картах ниже). На самом деле данные ДНК-генеалогии показывают, что носители гаплогруппы R1b двигались тысячелетиями в противоположном направлении – от Южной Сибири на запад, создав на этом пути хвалынскую и ямную культуры. Обратного движения эрбинов, носителей гаплогруппы R1b, на восток уже не было. Нет их ни в Индии (R1b там вообще практически нет), ни на Урале, ни в Зауралье с датировками, более поздними, чем в ямной культуре, как должно было бы быть, если движение от ямной культуры было на восток. Причина таких заблуждений популяционных генетиков уже была показана в предыдущих частях этой серии – у них системные проблемы и ошибки в методологии геномного анализа. Вся методология базируется на сравнительном сопоставлении некоего диффузного поля сотен тысяч и миллионов нуклеотидов с мутациями и без, которые подразделены на множество фрагментов разной длины. В итоге хрестоматийный вопрос «где талию будем делать» отдается на полный откуп компьютеру, применяя множество программ типа «admixture f3 statistics». А поскольку компьютеру не поясняют, что гаплогруппа R1a из гаплогруппы R1b не происходит (как и наоборот, как не происходит и одна «параллельная» гаплогруппа из другой), такие ограничения в программу популяционные генетики не вводят, вот компьютер и принимает важнейшие и несовместимые снипы за совершенно совместимые, сваливая и те, и другие в общую кучу. Результат – налицо.
http://pereformat.ru/wp-content/uploads/2016/08/r1a-migration32.jpg
Карта показывает как неверное направление, так и неверную концепцию, что культура шнуровой керамики (R1a) якобы произошла от ямной культуры (R1b), и что ямная культура принесла на запад индоевропейские языки (так называемая «степная гипотеза»). Эта принципиальная ошибка – результат недавних геномных исследований, методология которых основывается на «сопоставлении» фрагментов геномов и поиске «похожестей» совокупностей фрагментов, которых по сути дела существует практически бесконечное множество. Компьютерная программа, следуя заложенным инструкциям, сама решает, что на что более похоже. Это уже привело к множеству жалоб ДНК-тестируемых на интерпретацию или личных данных, в том, что программы несовершенны, и постоянно ошибаются, потому что критерии выбора совершенно расплывчаты. Подробнее – в тексте. Карта – из статьи (Allentoft et al., 2015).
http://pereformat.ru/wp-content/uploads/2016/08/r1a-migration33.jpg
Карта показывает как неверное направление, так и неверную концепцию, что афанасьевская культура якобы произошла от ямной культуры (R1b). Эта принципиальная ошибка – результат недавних геномных исследований, см. пояснение в подписи к предыдущей карте и описания в тексте. Карта – из статьи (Allentoft et al., 2015).
Были рассмотрены ископаемые ДНК пяти образцов афанасьевской культуры (Allentoft et al., 2015), но все они оказались женскими, и дали следующие митохондриальные ДНК: U5a1a1 (дважды), T2c1a2 и J2a2a (дважды). Датировки, соответственно, следующие: 5140±200, 4800±100 и 4750±220 лет назад. Как видим, они довольно близки друг другу, и в целом примерно соответствуют приведенному выше возрасту афанасьевской культуры как 5200-4500 лет назад, так и в целом III-II тыс. до н.э., принимаемому в последнее время историками.
Митохондриальные ДНК обычно малоинформативны, за исключением немногих специальных вопросов. Причина довольно проста – женщины обычно приходят в селение к мужу, не наоборот. В качестве примера можно привести нашу фамильную деревню в Курской области – на протяжении столетий в деревне была одна мужская гаплогруппа – R1a, точнее (с субкладами) R1a-Z465-Z280-CTS1211-CTS3402-CTS8816-Y2902-YP1447. А митохондриальных ДНК были наверняка десятки, а то и сотни, из многих курских деревень, в основном деревень однодворцев, откуда приходили и куда уходили невесты и жены, а то и дальше, когда жен привозили из далеких военных походов. Еще пример – из гарема выходит один субклад мужской гаплогруппы, но десятки разных мтДНК. Поэтому заниматься ДНК-генеалогией с помощью мтДНК – в большинстве случаев дело заведомо проигранное. И действительно, если мы посмотрим, какие еще ископаемые ДНК имеют мтДНК U5a1a1, то таких в списке (Allentoft et al., 2015) оказывается 26. Но мы отберем (см. ниже) из них только те, которые старше афанасьевских, рассчитывая на то, что есть вероятность, что носители соответствующих гаплогрупп пришли в афанасьевскую культуру (или ее создали) со своими женщинами той же гаплогруппы, которую они унаследовали от своих матерей, бабушек, прабабушек, и далее по линии женской мтДНК. У мужчин мтДНК по наследству не передается, поэтому мужская ДНК-генеалогия здесь бессильна. Сразу заметим, что это предположение оказалось неверным, ямники в афанасьевскую культуру не приходили, и афанасьевскую культуру не создавали.
Важное замечание – если носители гаплогруппы R1a пришли на Русскую равнину из Европы только примерно 4500 лет назад, то как они могут оказаться в Афанасьевской культуре 4700-5100 лет назад (как датируются афанасьевские мтДНК)? Та же ведь на многие тысячи километров восточнее, туда еще дойти надо. Иначе говоря, если R1a в афанасьевской культуре будут действительно европейскими, то это должны быть какие-то особые европейские субклады, старше, чем R1a-Z645, которых пока не находили по пути арийских миграций. Но поскольку в афанасьевской культуре пока никаких Y-хромосомных субкладов не нашли, то дискуссия в таким ключе бессмысленна. Мы только знаем, что ископаемых R1a с археологическими датировками старше 4700-5100 лет назад за Уралом не находили, все были не старше 4100 лет. Если на Алтае, в Монголии или Китае окажутся ископаемые носители R1a (или расчетные общие предки) старше 4700 тысяч лет, то это должны быть автохтоны, и у них вряд ли окажутся субклады R1a-Z93 или R1a-Z280.
Еще одно важное замечание – если мтДНК совпали в Сибири, и, скажем, в Европе, то опять есть немало вариантов объяснений такого совпадения. Во-первых, направление маршрута могло быть в любую сторону – из Сибири в Европу или из Европы в Сибирь. Или из некой средней «точки» в обе стороны. В какую именно сторону оно было, надо разбираться по дополнительным факторам. Во-вторых, это совпадение может быть не напрямую, а через унаследование от общих предков, которые могли жить тысячелетия назад. Например, «байкальский мальчик» с датировкой 24 тыс. лет назад имел мтДНК U. От его родственников по женской линии могли пойти потомки и в Сибирь, и независимо в Европу, поэтому совпадение мтДНК в Сибири и в Европе на самом деле мало что значит (в отношении направления миграции), даже если в Европе датировка мтДНК окажется старше, чем в Сибири. А компьютер может выбрать любой вариант направления миграции.
Но тем не менее, для образовательных целей посмотрим, что получается. Итак, афанасьевскую мтДНК U5a1a1 или более старшую, как, например, U5a1а или U5a1, и с возрастом более 4700 лет, имели следущие ископаемые ДНК:
— U5a1, Швеция, датировка 7730±180 лет назад, мужчина, гаплогруппа I2c,
— U5a1a1, ямная культура (Самарская область), датировка 4840±190 лет назад, мужчина, гаплогруппа R1b-Z2105,
— U5a1a1, ямная культура (Самарская область), датировка 5130±190 лет назад, мужчина, гаплогруппа R1b-M269.
C мтДНК T2c1a2:
— T2c1, женщина, Германия, датировка 7120±100 лет назад,
— T2c1a2, мужчина, ямная культура (Самарская область), датировка 5160±230 лет назад, гаплогруппа R1b-Z2105.
C мтДНК J2a2a:
— J2a2a, мужчина, Германия, датировка 4290±225 лет назад, гаплогруппа P1-P228,
Последний случай за неимением других примеров цитирует датировку даже более позднюю, чем в Афанасьевской культуре, хотя субклад мтДНК тот же.
Как и ожидалось, мтДНК нам не помогли. Но стало понятнее, почему компьютер направил исследователей по ложному пути, и «выдвинул» вариант, что ямники отправились в афанасьевскую культуру, хотя это не проходит по другим вариантам. МтДНК, например, оказались сходными, как, видимо, и ряд фрагментов ДНК в ямной и афанасьевской культуре. Поскольку датировки примерно одинаковые в ямной культуре и афанасьевской, и ясно, что прямого контакта быть не могло, между ними тысячи километров, у них просто один мтДНК-общий предок, но компьютер в таких деталях не разбирается. Похожи – значит похожи. Это компьютер и выдал. А попгенетики посмотрели на концепции историков, и увидели, что ими предлагался вариант, по которому афанасьевскую культуру «основали ямники». Всё, ответ получен, вот и компьютер подтвердил. Так популяционные генетики «делают науку». Они же не могут сказать, что историки ошибаются, это было бы некошерно, да и вызвало бы подозрение, как они там считают. Зачем это им нужно?
В итоге мы пока не знаем, какие гаплогруппы преобладают в афанасьевской культуре, какие там субклады и какие датировки общих предков. А соответствующие «геномные данные» следует просто отбросить за профнепригодностью.
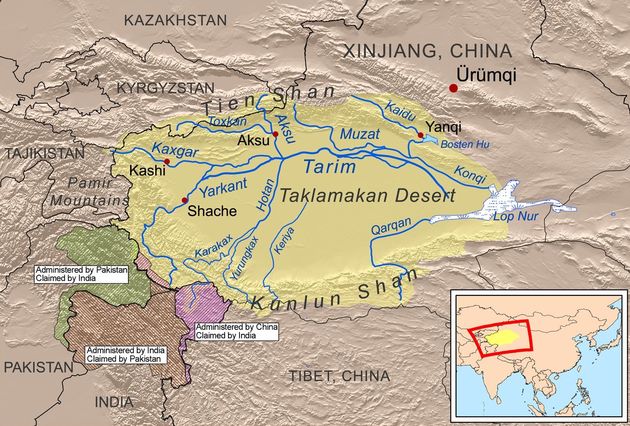
Карта ниже показывает общий вид географии Таримского бассейна и пустыни Такла-Макан, в которой были найдены «таримские мумии», с археологической датировкой 4000±40 лет назад. Все тестированные поначалу семь человек показали гаплогруппу R1a, гаплотипы не определяли (или их не сообщили). Более недавнее их тестирование китайскими исследователями показало одиннадцать R1a, и одну гаплогруппу К. По предварительным данным, ископаемые ДНК не показали субклад Z93, то есть это не арии и не их потомки. Более того, таримский гаплотип гаплогруппы R1a показал такие экзотические мутации, причем много, что общий предок его с гаплотипами R1a Русской равнины жил не менее 23 тысяч лет назад. Случайно или нет, но это практически датировка образования гаплогруппы R1a (20-22 тысяч лет назад), определенная и по мутациям в гаплотипах, и по снипам.
http://pereformat.ru/wp-content/uploads/2016/08/r1a-migration34.jpg
Общий вид карты Таримского бассейна в пустыне Такламакан, где были найдены «таримские мумии» с гаплогруппой R1a и археологической датировкой 4000±40 лет назад.
В статье (Li et al., 2010), в которой описана находка мумий в Таримском бассейне, рассуждают достаточно распространенным, но увы, некорректным образом. Например, если были обнаружены антропологические европеоиды, то они непременно являются мигрантами из Европы. Если там гаплогруппа R1a, то она непременно из Европы. Если они одеты в ткани, похожие на шотландский плед, то их выход состоялся непременно из Шотландии. Обратный вариант авторами не рассматривается и совершенно напрасно. Как пояснялось выше, мы не можем исключить наличие там автохтонных R1a, тем более, если там действительно не оказалось субклада Z93, распространенного в Центральной Азии. Тип древних тканей мог бы быть принесен не из Европы, а наоборот, в Европу. Однако когда я высказал свое сомнение Виктору Меиру, основному автору статьи про гаплогруппу R1a таримских мумий (гаплотипы они не определяли или не публиковали) про направления миграций древних таримцев, если их полностью основывать на виде тканей, которые выглядят как европейские (согласно д-ру Меиру), то он впал в раздражение и посоветовал мне читать его книги про европейские ткани. Он был уверен, что если гаплогруппа R1a – то непременно из Европы. На этом наша дискуссия завершилась.
Да, таримские носители R1a могли прибыть из Европы 4000 лет назад или ранее, но это необходимо доказывать, а в отсутствие гаплотипов это практически безнадежно. Почему авторы статьи не показали или не определили гаплотипы — остается загадкой. Авторы исследования (Li et al., 2010) предполагают, что обнаруженные люди антропологически близки к носителям афанасьевской археологической культуры, что еще более смещает в древность обнаруженных носителей R1a, и делает сомнительным их прибытие из Европы 4000 лет назад или ранее. Но это просто предположение, поскольку европеоиды – это далеко не только афанасьевцы. Таким образом, альтернатива остается нерешенной – либо это древние, автохтонные R1a, европеоиды по происхождению, хотя в Европе никогда не были, как и их предки, либо это потомки мигрантов, которые вышли из Европы не позднее 5-6 тысяч лет назад и пересекли Евразию значительно раньше, чем «классические» арии гаплогруппы R1a-Z645-Z93.
Последнее, в принципе, не исключено, но пока нет достаточного количества фактов, подтверждающих предположение о двух раздельных миграциях ариев через всю Евразию с разницей в полторы-две тысячи лет, то есть с прибытием на Алтай 5500 и 3500 лет назад. Данные факты могли бы также перевернуть представления о том, кем были создатели цивилизации Хараппы и Мохенджо-Даро 5 тысяч лет назад, и как там оказалась арийская свастика в те времена.
Еще одной проблемой с таримскими мумиями является их отнесение к тохарам, то есть предкам людей, говоривших на тохарских языках (которые считаются наиболее восточными индоевропейскими языками), и предположительно проживавших 1700-1200 лет назад, то есть уже в нашей эре. Тохар помещают в Таримский бассейн, им приписывают обнаруженные в начале 20-го века рукописи, которые датируют 6-8 вв н.э., и язык (ныне мертвый) подразделяют на два – «тохарский А» (восточно-тохарский) и «тохарский В» (западно-тохарский). Предполагают, что источник тохар – афанасьевская культура (что, конечно, вообще ни на чем не основано, очередная фантазия), предполагают, что тохары пришли из Европы, когда – неизвестно. В общем, видно, что все связи между людьми, языками, предполагаемыми источниками и их датировками – сугубо условные. Неизвестно и то, являются ли таримские мумии тохарами. Это всё – рассуждения «по понятиям». Всё, что мы знаем, это то, что тохарские мумии с датировкой примерно 4000 лет назад имели гаплогруппу R1a, публикаций о субкладе нет, но есть предварительные данные, распространенные китайскими генетиками, что они не Z93, и что гаплотипы (на одном примере) очень необычные, и – по нашим данным – если определены правильно, то их общий предок жил примерно 23 тысяч лет назад. Это – всё.
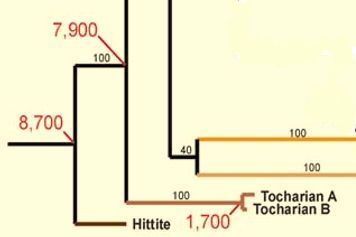
Есть еще одна нестыковка в предположениях о тохарах. Их язык, тохарский, отправляют на глубину 7900 лет назад.
Фрагмент лингвистического дерева, включающий тохарские языки с датировкой 7900 лет назад (по данным Gray and Atkinson, 2003).
Ситуация подобна той же с хетскими языками, о чем мы говорили в предыдущей части этой серии. Либо (1) лингвистические расчеты неверны (что вполне возможно, поскольку зависят от ряда постулатов, приближений и допущений), либо (2) тохары не выходили из Европы, и ПИЕ (прото-индоевропейские) языки – автохтонные в Южной Сибири, и оттуда принесены через Анатолию и Балканы в Европу, либо (3) они как древняя ветвь гаплогруппы R1a вышли из Европы и вернулись в Южную Сибирь немедленно после прибытия в Европу 10-8 тысяч лет назад. Насколько автору настоящей статьи известно, ни одна из этих гипотез не рассматривалась или во всяком случае серьезно не анализировалась. Поначалу распространение получила точка зрения, что тохарские языки – кентумные, и что тохары, соответственно, имели гаплогруппу R1b. Эта точка зрения, как принято у популяционных генетиков, скоро приобрела статус факта, и была непререкаемой много лет, но потом было найдено, что таримские мумии имеют гаплогруппу R1a, и хотя никто не показал, что они именно тохары, «факт» про гаплогруппу R1b моментально умер, и больше его никто не вспоминает. Тоже характерно для популяционных генетиков.
А что еще рассматривалось? Много места в своих рассуждениях посвятил тохарам Л.С. Клейн. В его книге «Древние миграции и происхождение индоевропейских народов» (2007, не издана, доступна в сети) тохары упоминаются почти 200 раз. И какой «сухой остаток»? Да, в общем, его нет. Есть серия предположений, что (цитаты):
– Предковые диалекты тохар пришли на восток из центра Европы…
– Несомненная дальняя миграция тохаров на восток из участка… в ареале праиндоевропейцев…
– арии и тохары ушли дальше (от Европы)…
– факт дальней миграции (тохар) бронзового века здесь несомненен.
– не позднее III в. до н.э. …тохары… жили по соседству с Китаем и Индией, т.е. скорее всего уже в Синьцзяне, на Тариме.
– предположение об афанасьевской культуре как тохарах не находит подтверждения.
– допустимо… предположить, что карасукская культура – это тохары…
– тохары двигались из Европы на восток по лесной полосе далеко от степей (ареала ариев) и долго жили в финноугорской среде.
– Видимо, исходный пункт движения прототохаров на восток лежал в восточной части и на севере Центральной Европы.
– Формирование фатьяновской культуры, видимо, представляет собой материальную основу выделения тохарских языков из индоевропейской общности или завершения этого процесса…
– прототохарский язык – это пришлый индоевропейский язык, усвоенный местным неиндоевропейским населением…
– Неясно, где проходило усвоение индоевропейского языка иным населением – уже в местах обнаружения тохарских языков или где-то поближе к праиндоевроейскому.
– нивхи в прошлом населяли большую территорию на Дальнем Востоке – Приамурье и Сахалин и вполне могли быть тем этносом, который послужил субстратом для тохарского
– моя фатьяновско-карасукская концепция происхождения тохаров…
– Тохары двигались южнее, где ассимилировали население с каким-то агглютинирующим языком, возможно, уральской семьи, продвинувшееся на юг со своей прародины… Таким же субстратом для тохаров мог быть и дравидский…
– ни одного заимствования дравидского в тохарских или тохарского в дравидском нет.
Как видно, ничего определенного. Показательно, что Клейн выдвигал «гипотезы», проверить которые тогда, несколько лет назад, казалось невозможным, и в этом была их прелесть. Прошло несколько лет, и появилась ДНК-генеалогия, которая повергла Клейна в величайшее раздражение и скорбь (см. книгу: Клёсов, А.А. Кому мешает ДНК-генеалогия. М., Книжный мир, 2016 г., 848 стр.). И понятно, почему.
Видимо, с тохарами круг остается замкнутым. То, что таримские мумии показали R1a, не значит, что они были тохарами. А нахождение любых костных останков для ДНК-анализа встретит проблему идентификации, что они были именно тохарами. Лингвисты и археологи пока не могли выйти из этого круга, а без них ДНК-генеалогия не сможет привязать ископаемые ДНК к тохарам. Из этого круга надо выходить, но пока непонятно, как.
Из археологических данных известно, что среди скифов были монголоиды. Но надо сказать, что если бы у тех монголоидов были определены гаплогруппы, то с хорошей вероятностью у них тоже была бы гаплогруппа R1a. Не только одна R1a, но определенно в заметной степени. Как такое может быть? И вот здесь – новый виток сведений об алтайских скифах, потомках ариев. Переходим к пазырыкской археологической культуре и современным жителям Алтая с гаплогруппой R1a. Пазырыкская культура – это археологическая культура железного века (III-V вв. до н.э., хотя некоторые опускают дату до VI века до н.э.), которую относят к «восточному скифскому кругу». Регион – горный Алтай и смежные территории Алтая, Казахстана и Монголии. Основное занятие – кочевое скотоводство. Высказывается предположение, что пазырыкская культура – производная от афанасьевской.
Недавно изучались гаплотипы и гаплогруппы (мужские и женские) современных жителей этого региона (Dulik et al., 2012), и митохондриальные гаплогруппы (женские по сути, поскольку мужчины получают их от матери, но дальше не передают, нет митохондрий в сперматозоидах) ископаемых костных останков пазырыкской культуры (Gonzalez-Ruiz et al., 2012). Оказалось, что большинство современных мужских гаплогрупп в регионе относятся к гаплогруппе R1a, причем наибольшее их количество было в народности алтай-кижи. R1a содержали также тубалары, челканы, кумандины. На втором месте по численности была гаплогруппа Q, затем С, потом N, остальные – минорные, единичные гаплогруппы, включая R1b, которые, как правило, случайны, и могли попасть туда в любое время.
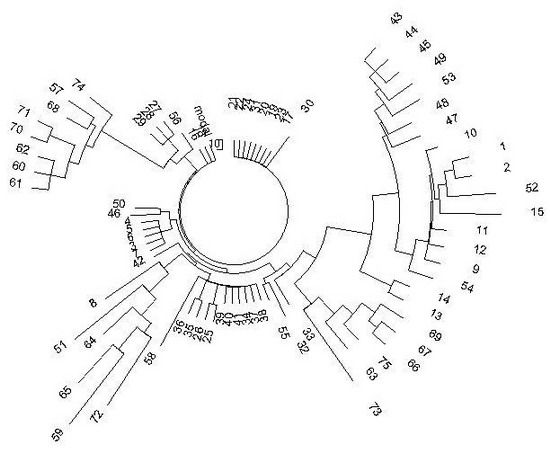
Однако у носителей гаплогруппы R1a на Алтае была характерная особенность. Если на Русской равнине и в Центральной Европе у них преимущественно митохондриальная гаплогруппа (мтДНК) Н, как и у их жен и подруг, так называемая «европейская», или «западная» мтДНК, то на Алтае у R1a, в основном, восточные, «восточно-евразийские», азиатские мтДНК – A, C, D и G, их у носителей гаплогруппы R1a до половины и двух третей, остальные – минорные, единичные. Европеоидных мтДНК у них почти нет. Носители мтДНК A, C, D и G, как правило – монголоидные женщины и их сыновья и дочери, опять же монголоидные. Вот и разгадка, почему алтайские носители гаплогруппы R1a, потомки ариев-скифов, как правило, сами монголоидны. Антропологию в значительной степени определяют женщины. К тому же, у алтайских R1a те же монголоидные женщины, видимо, сменили язык всей популяции на тюркский. Анализ гаплотипов алтайцев с точки зрения ДНК-генеалогии проведен в статье (Клёсов, 2012). Дерево гаплотипов имеет необычный вид:
Дерево из 75 гаплотипов гаплогруппы R1a на Алтае в 17-маркерном формате. Верхняя левая ветвь – тубалары (57, 68, 70, 71, 74), челканы (60, 61, 62) и алтай-кижи (57). Нижняя правая ветвь – подобный состав: тубалары (66, 67, 69, 73, 75) и один челкан (63). Нижняя левая ветвь – смешанная: кумандины (64, 65), тубалар (72), алтай-кижи (8, 51, 59). Остальные гаплотипы – алтай-кижи. Гаплотипы приведены в статье (Клёсов, 2012).
Вид необычный потому, что гаплогруппа одна, то есть род один, а ветви расходятся в разные стороны, будучи заметно изолированными друг от друга. Это означает, что у данной популяции была тяжелая судьба. Племена погибали, немногие спасшиеся бежали, начинали свои генеалогические линии практически с нуля. Это повторялось, и опять спасались, бежали, и опять начинали свои линии. То есть повторялся эффект «последнего из могикан». В итоге все ветви на дереве гаплотипов относительно молодые, это набор относительно молодых «кустов», пересаженных со старых черенков, которые погибли. Но исходный общий предок жил в далекой древности, на это указывают масштабные расхождения между ветвями. Для сравнения, дерево гаплотипов группы R1a на Русской равнине в том же 17-маркерном формате выглядит несравненно более благополучно.
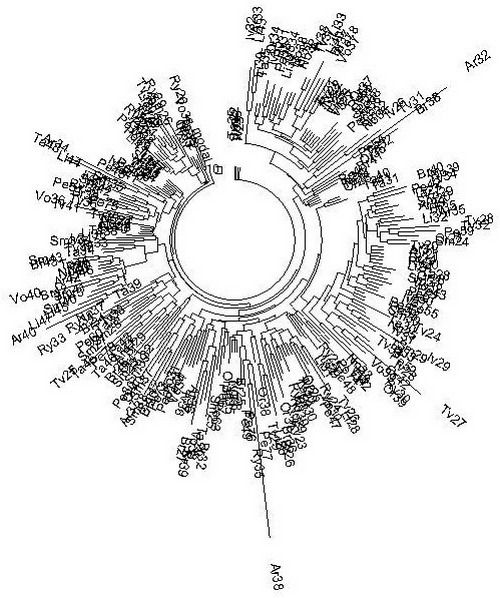
Дерево из 257 гаплотипов гаплогруппы R1a на Русской равнине (по 12 областям Российской Федерации) в 17-маркерном формате. Резко выделяющиеся гаплотипы Ar32 и Ar38 (из Архангельска) попали в список ошибочно, они относятся к другим гаплогруппам. Они оставлены на дереве, чтобы показать, насколько дерево чувствительно к «чужакам» (Клёсов, 2009).
Еще более благополучно выглядит дерево гаплотипов Русской равнины для большого числа протяженных, 67-маркерных гаплотипов.
Дерево из 801 гаплотипов гаплогруппы R1a на Русской равнине – в 67-маркерном формате (Rozhanskii and Klyosov, 2012).
Посмотрим более внимательно на современные алтайские гаплотипы. Левая верхняя ветвь из восьми гаплотипов (тубалары и челканы) имеет следующий предковый гаплотип:
13 24 16 9 12 14 10 14 11 32 – 14 14 20 12 17 11 23 (алтайские гаплотипы, ветвь)
В ветви всего 10 мутаций на 8 гаплотипов, то есть на 8х17 = 136 маркеров, что дает 10/8/0.0365 = 34 → 35 условных поколений, или 875±290 лет до общего предка (стрелка – поправка на возвратные мутации, 0.0365 – константа скорости мутации для 17-маркерных гаплотипов). Иначе говоря, общий предок этой ветви жил примерно в 12-м веке нашей эры, плюс-минус три века. Ясно, что он удален от скифов по времени, но гаплотип показывает, что современные алтайцы – прямые потомки ископаемых андроновцев, с археологической датировкой 3800-3400 лет назад. Тип гаплотипа тот же, та же самая пара 14-32.
13 25 16 11 11 14 10 14 11 32 – 15 14 20 12 16 11 23 (ископаемые андроновцы, R1a)
Между ними – шесть мутаций, что разводит ископаемых андроновцев и общего предка современных алтайцев на 6/0.0365 = 164 → 196 условных поколений, или примерно на 4900 лет, и помещает общего предка современных алтайцев и ископаемых андроновцев на (4900+3600+875)/2 = 4700 лет. Это и есть арии на Русской равнине. Иначе говоря, разница в шесть мутаций практически точно соответствует тому, что современные алтайцы – потомки древних андроновцев, а те – древних ариев.
Те же расчетные операции по всем ветвям дерева алтайских гаплотипов привели к тому же результату – все ветви происходят от древних андроновцев, но сами ветви – обрывки единого массива потенциальных потомков, которые не выжили, выжили только «лохмотья» от ветвей древних андроновцев, выжила серия мини-популяций с общими предками от 875 до 750 лет назад, датировки которых равны друг другу в пределах погрешностей расчетов. Основной вывод – что эти современные алтайские гаплотипы, точнее, их носители – потомки древних скифов, в свою очередь, потомков древних ариев, которые жили на Русской равнине.
Раскопки пазырыкской культуры выявили три мтДНК, относящиеся к бронзовому веку и шестнадцать мтДНК – к железному веку. К сожалению, Y-хромосомные ДНК не изучались, но мы уже знаем, что они с большой вероятностью дали бы, в основном, гаплогруппу R1a. Но и полученная информация оказалась важной. Из всех 19 ископаемых мтДНК гаплогрупп 11 оказались азиатскими (A, C, D, и G), и 8 – западными, точнее, западно-евразийскими (HV, J, U, T, K). Все три гаплогруппы бронзового века оказались азиатскими. Гаплогруппы железного века дали смесь европейских и азиатских гаплогрупп. Это в любом случае показывает, что скифы были и европеоидные, и монголоидные, причем арии-скифы, пришедшие в алтайский регион в бронзовом веке, то есть самые ранние, брали в жены местных монголоидных женщин, и их потомки, сохраняя гаплогруппу R1a, были уже монголоидными. Это опять объясняет монголоидность некоторых (или многих) скифов, кочевавших по степям Евразии. Но многие скифы, очевидно, откочевывали на восток со своими европейскими женами и подругами, что и дало «западные» мтДНК в их потомках, в том числе и на Алтае в древности. Вот и получалось разнообразие антропологии скифов-кочевников, от европеоидных до монголоидных, при наличии основной гаплогруппы R1a.
На самом деле древние арии на Русской равнине и были предками славян гаплогруппы R1a, если судить по пантеону (языческих) богов, которые перекликаются от Восточной Европы через Русскую равнину до Индостана, а также сказаниям и мифам. Их топонимы и гидронимы были древними славянскими просто по определению. И не нужно сравнивать их звучания с современными славянскими, язык с тех пор изменился, и не он должен лежать в основе классификации древних племен и народов. А вот характерные «подписи» в их ДНК не изменились, и унаследованы без принципиальных изменений в ходе многих тысячелетий и десятков тысяч лет, только естественным путем разветвились, оставляя общую картину доступной для несложной реконструкции. Языки здесь – вторичный признак, не главный, они изменчивы и принципиально подвержены произвольным толкованиям и интерпретациям лингвистов. Которые, кстати, и сами между собой не могут договориться в подавляющем большинстве случаев. И если мы по всей справедливости будем считать языки в данном контексте вторичными факторами, то картина вырисовывается вполне четкая: арии, скифы и восточные славяне – это одни и те же люди, в своей естественной хронологической динамике, которая начала расходиться на ветви от субклада R1a-Z645 пять с лишним тысяч лет назад. Большинство их принадлежало и принадлежит одному и тому же роду – R1a.
Теперь вопрос – а насколько «большинство»? Полагаю, точная цифра здесь не имеет значения. Ясно, что в их составе были и другие гаплогруппы, но они не доминировали. Во всяком случае, данных таких нет, будут – рассмотрим. У гаплогруппы R1b была своя интересная история, но в составе ариев, прибывших в Индию, их не было. Во всяком случае, в современной Индии их очень мало, в высших кастах практически нет, и среди 367 браминов, тестированных на гаплогруппы, не было найдено ни одного случая гаплогруппы R1b (Sharma et al., 2009). Гаплогруппу R1a имели до 72% высшей касты в Индии, в основном среди браминов. Среди современных этнических русских носителей гаплогруппы R1b примерно 5%. Для сравнения, гаплогруппы R1a среди этнических русских – до двух третей в южных областях – Курской, Белгородской, Орловской. В среднем, по всем европейским областям Российской Федерации, включая северные (во многом финно-угорские) – половина R1a от всего населения.
Еще соображение, вытекающее из сказанного выше. Все известные деления скифов на «царских скифов», «скифов-пахарей», «скифов-воинов», «скифов-кочевников», «скифов-земледельцев», «борисфенитов» (Геродот) и прочих являются поверхностными. По подобным «признакам» мы и сегодняшних россиян можем делить на «пахарей», «воинов», «инженеров», «профессоров», «медицинских работников» и прочих, но относится ли это деление к происхождению русского народа? Хотя делить и изучать можно и нужно, и этим занимаются социальные службы, но надо понимать, с какими целями, для чего это стоит делать, и на какие загадки истории – в случае скифов – это ответит.
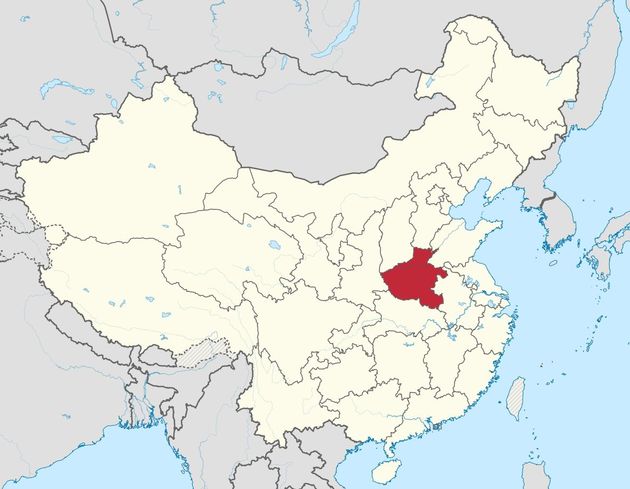
В первой части этой серии статей мы рассмотрели удивительные данные о сходстве керамики трипольской культуры (7500-4650 лет назад) и китайской культуры Яншао (8000-4000 лет назад). Эти данные наводят на мысль о взаимосвязи этих народов в древности. Поэтому не стало большой неожиданностью, что довольно большая серия гаплотипов гаплогруппы R1a была выявлена у современного населения китайской провинции Хэнань (Meisen et al., 2015), на востоке центральной части Китая. Как видно на приведенной ниже карте, этот регион весьма удален от Индии, от северо-западной китайской территории Синцьзянь, от Сибирского Алтая (на стыке четырех регионов в верхнем левом секторе карты, от Южного Урала, и тем более от степной и лесостепной полосы юга Украина и России, но хэнаньские гаплотипы оказались очень похожими на гаплотипы этнических русских гаплогруппы R1a. Это было показано построением дерева из 76 гаплотипов китайских гаплотипов и разделением их ветвей, как показано в статье И.Л. Рожанского на Переформате.
Китай и провинция Хэнань
http://pereformat.ru/wp-content/uploads/2016/08/r1a-migration42.jpg
Республика Алтай Российской Федерации
Поскольку китайские исследователи предпочли нестандартный 22-маркерный формат гаплотипов, в обычной ситуации сопоставить гаплотипы ветвей с другими известными гаплотипами было затруднительно. Но мы работаем с 111-маркерными гаплотипами, где все эти необычные маркеры представлены. В процитированой статье И.Л. Рожанский привел базовые гаплотипы трех основных ветвей дерева китайских гаплотипов, а также оставшихся более древних гаплотипов, которые остались вне ветвей, и рассчитал, когда жили их общие предки:
13 25 16 11 11 14 12 10 13 11 17 – 16 25 14 21 33 – 12 16 11 – 14 10 23 (550±140 л.н.)
13 25 16 10 11 15 12 11 13 11 19 – 16 24 14 19 32 – 13 16 11 – 14 10 23 (325±80 л.н.)
13 23 15 10 11 14 12 10 12 11 17 – 15 24 14 19 30 – 12 15 11 – 12 10 24 (750±390 л.н.)
13 25 16 11 11 14 12 10 14 11 17 – 15 24 14 20 33 – 13 15 11 – 13 10 23 (2550±670 л.н.)
Приведем для рассмотрения индийские гаплотипы (R1a-Z645-Z93) в том же формате:
13 25 16 11 11 14 12 10 13 11 17 – 16 24 14 20 32 – 12 15 11 – 14 10 23
И гаплотипы этнических русских (R1a-Z645-Z280) в том же формате:
13 25 16 11 11 14 12 10 13 11 17 – 15 24 14 20 32 – 12 16 11 – 14 10 23
Мы видим, что индийские и русские предковые гаплотипы опять близки друг к другу, как было показано ранее в этой серии статей, между ними на вид две мутации (отмечены), а на самом деле там всего (0.83+0.66) = 1.49 мутаций, поскольку по всей серии гаплотипов мутации являются дробными, и разница между гаплотипами примерно равна 1.49/0.05 = 30 → 31 условное поколение, то есть 775 лет (0.05 мутаций на гаплотип за 25 лет – это константа скорости мутации для данных 22-маркерных гаплотипов). Мы видели, что по 37-маркерным гаплотипам эта разница составляла 1025 лет, что принципиально одно и то же, поскольку общие предки этих серий гаплотипов жили около 5 тысяч лет назад. По 37-маркерным гаплотипам их общий предок жил (4900+4600+1025)/2 = 5300 лет назад, по 22-маркерным – (4900+4600+775)/2 = 5140 лет назад. Это все в пределах погрешности расчетов.
Вернемся к китайским гаплотипам. Они различаются по мутациям от предкового гаплотипа этнических русских всего на 4, 8 и 5 мутаций, не считая совсем малой и неточной ветви из четырех гаплотипов, что помещает их общих предков с русскими гаплотипами на 4600±500 лет назад, то есть опять в Европу или немного восточнее, на Русскую равнину. От индийских гаплотипов китайские отличаются ровно на такое же количество мутаций, то есть их общий предок опять уходит в 4600±500 лет назад, опять в Европу или на Русскую равнину (хотя, строго говоря, Русская равнина это часть Европы, но понятно, о чем здесь речь). Получается, что китайские R1a – не из Индии, а с севера, с Русской равнины.
Надо сказать, что в провинции Хэнань сейчас живут 94 млн человек, и если считать, что выборка в 1100 гаплотипов, в которой 76 гаплотипов оказались R1a (6.9% от всех), отражает состав населения в Хэнане, и что половина населения – мужчины, то только в одной этой провинции живут более трех миллионов носителей гаплогруппы R1a. Это провинция считается самой большой колыбелью китайской культуры, первые городища там датируются 7 тысяч лет назад, знаменитая культура Яншао – 6000±2000 лет назад, но именно в этой культуре найдены характерные керамические изделия трипольской культуры (Klyosov and Mirinova, 2013). Поскольку культура Яншао датируется 8000-4000 лет назад, то, возможно, носители R1a принесли туда свою керамику (или искусство ее изготовления) почти в те же времена, как они появились на Южном Урале, в Синташтинской археологической культуре.
Продолжение следует…
Клёсов, А.А. (2009) Гаплотипы восточных славян: девять племен? Вестник Российской Академии ДНК-генеалогии, т. 2, № 2, 232-251.
Клёсов, А.А. (2012) Гаплотипы группы R1a на Алтае: «автохтоны» и «индоевропейцы». Вестник Академии ДНК-генеалогии, т. 5, № 12, 1511-1525.
Клёсов, А.А. (2016) Кому мешает ДНК-генеалогия. «Книжный мир», М., 2016, 848 стр.
Allentoft, M.E., Sikora, M., Sjögren, K.G., Rasmussen, S., Rasmussen, M., Stenderup, J., Damgaard, P.B., Schroeder, H., Ahlström, T., Vinner, L., Malaspinas, A.S., Margaryan, A., Higham, T., Chivall, D., Lynnerup, N., Harvig, L, Baron, J., Della Casa, P., Dąbrowski, P., Duffy, P.R., Ebel, A.V., Epimakhov, A., Frei, K., Furmanek, M., Gralak, T., et. al. (2015) Population genomics of Bronze Age Eurasia. Nature 522(7555), 167-72. doi: 10.1038/nature14507.
Dulik, M.C., Zhadanov, S. I., Osipova, L.P., Askapuli, A., Gau, L., Gokcumen, O., Rubinstein, S., Schurr, T.G. (2012) Mitochondrial DNA and Y chromosome variation provides evidence for a recent common ancestry between Native Americans and indigenous Altaians. Amer. J. Human Genetics, 90, 1-18. DOI 10.1016/ajhg.2011.12.014.
Gonzalez-Ruiz, M., Santos, C., Jordana, X., Simon, M., Lalueza-Fox, C., Gigli, E., Aluja, M., Malgosa, A. (2012) Tracing the origin of the East-West population admixture in the Altai region (Central Asia). PLOS One, 7, 1-11. e48904.
Gray, R.D., Atkinson, Q.D. (2003) Language-tree divergence times support the Anatolian theory of Indo-European origin. Nature 426, 435-439.
Hedjazi, A.A.T.A., Kerachian, M.A., Honarvar, Z., Dadgarmoghaddam, M., Raoofian, R. (2014) Genetic profile of 17 Y-chromosome STR haplotypes in East of Iran. Forensic Science International: Genetics, 14, doi:10.1016/j.fsigen.2014.10.010, e6-e7.
Keyser, C., Bouakaze, C., Crubezy, E., Nokolaev, V.G., Montagnon, D., Reis, T., Ludes, B. (2009) Ancient DNA provides new insights into the history of south Siberian Kurgan people. Human Genetics 126, 395-410.
Klyosov, A.A., Mironova, E.A. (2013). A DNA Genealogy Solution to the Puzzle of Ancient Look-Alike Ceramics across the World. Adv. Anthropology, Vol. 3, No. 3, pp. 164-172.
Li, C., Li, H., Cui, Y. et al. (2010) Evidence that a West-East admixed population lived in the Tarim Basin as early as the early Bronze Age. BMC Biology, 8:15, doi:10.1186/1741-7007-8-15.
Meisen Shi, M., Liu, Y., Zhang, J., Bai, R., Lv, X., Ma, S. (2015), Analysis of 24 Y chromosomal STR haplotypes in a Chinese Han population sample from Henan Province, Central China. FSI Genetics, 17, 83-86.
Rozhanskii, I.L., Klyosov, A.A. (2012) Haplogroup R1a, its subclades and branches in Europe during the last 9000 years. Adv. Anthropol. 2, No. 3, 139-156.
Sharma, S., Rai, E., Sharma, P., Jena, M., Singh, S., Darvishi, K., Bhat, A. K. et al. (2009). The Indian origin of paternal haplogroup R1a1* substantiates the autochthonous origin of Brahmins and the caste system. Journal of Human Genetics, 54, 47-55.
Анатолий А. Клёсов,
доктор химических наук, профессор
Перейти к авторской колонке





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}